 Ако предлагате частни уроци, може да рекламирате услугите си напълно безплатно!
Ако предлагате частни уроци, може да рекламирате услугите си напълно безплатно!
ПРОКАРИОТНА КЛЕТКА
При разглеждането на живите системи е логично да започнем от
най-простите. Термините "прост" и "сложен" се отнасят до степента на
организация. Колкото по-сложна (високо организирана) е дадена система, толкова е
по-подредена, т.е. по-малко вероятно е изграждащите я атоми да заемат местата си
случайно. Обикновено, макар и не винаги, по-сложните системи са и
по-големи.
Според типа на своите клетки организмите се делят на две
големи групи: прокариоти (гр. предядрени) и еукариоти (същинскоядрени). Както
показват имената им, решаващият разграничителен или "диагностичен" белег е
наличието или липсата на оформено ядро, но всъщност разликите са по-дълбоки.
Прокариотните клетки са по-прости от еукариотните, затова ще започнем с
тях.
С прокариотите следва да започнем и ако разглеждаме живия свят
исторически, т.е. от по-ранни към по-късни организми. Клетките в древните
пластове са прокариотни, за което се съди по малките им размери и простото им
устройство. Следователно съвременните прокариоти имат примитивни белези в
устройството си. Някои техни представители дори напомнят конкретни
микровкаменелости.
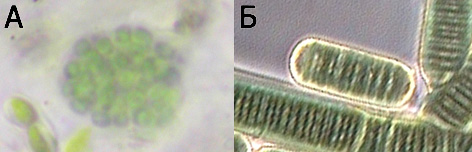
Живи цианобактерии, подбрани така, че да приличат
на показаните в предишния раздел микровкаменелости: А – неидентифицирана
колониална форма (клетките долу вляво са еукариотни водорасли), Б – Lyngbya sp.
Б е от Schauder (1997), култтивирана от J. Waterbury, с любезното разрешение на
авторите Rolf Schauder и David Graham.
Термините "примитивен" и "напреднал" също се използват за сравняване на видове, но за разлика от "прост" и "сложен" имат не организационен, а еволюционен смисъл. "Примитивен" значи подобен на нещо, възникнало рано в еволюцията. Ако, обратно, организмът в еволюцията си се е отдалечил от изходната форма, той се нарича напреднал, а белезите му – производни или апоморфни. В нашия курс често ще търсим примитивни белези у съвременни организми, но при това трябва да се внимава. Когато наричаме древните клетки прокариотни, имаме предвид само, че вкаменените им останки на вид са по-скоро прокариотни, отколкото еукариотни. Съвременните прокариоти са еволюирали точно толкова дълго, колкото и ние, и съвсем не всичко у тях е примитивно.
1. Разнообразие на прокариотите
Когато става дума
за прокариоти, най-напред се сещаме за разнородната група, наричана "бактерии".
Тя включва много причинители на болести, но не се изчерпва с тях. Прокариоти са
и гореспоменатите цианобактерии, които са фотоавтотрофи. Понякога те се наричат
синьозелени водорасли, но това име не е много подходящо, понеже истинските
водорасли са еукариоти. Трета група прокариоти са актиномицетите, чието тяло е
удължено и усложнено така, че външно напомня мицела на еукариотните гъби. Те
придават свойствения на почвата мирис и са важни производители на антибиотици.
Всички тези и някои други прокариоти се обединяват в една голяма група –
еубактерии или бактерии в широк смисъл. (Терминът "бактерия" понякога се
използва и като синоним на "прокариот" – трябва да се внимава за
подтекста.)
Освен еубактериите има и други прокариоти, наречени
архебактерии или само археи. Името намеква за древен произход, но истинската им
възраст не е ясна. Те са представени с малък брой видове в неблагоприятни
("екстремни") местообитания. Делят се на три групи: халобактерии, обитаващи
силно солени водоеми, метанобактерии или метаногенни бактерии, които живеят
главно в блатата и редуцират СО2 до СН4, и термоацидофилни бактерии, откривани в
горещи и богати на сяра води.
2. Устройство на клетката
Прокариотните клетки обикновено са дълги
1 – 10 микрометра. С малки изключения имат форма на кълбо, пръчица или извита
пръчица. Според формата си бактериите имат специални имена, например
кълбовидните се наричат коки.
Обикновено прокариотният организъм е
единична клетка. Понякога клетките-потомци от няколко деления остават свързани в
колония. При актиномицетите и много цианобактерии "колонията" е нишка от клетки,
между които има връзка и дори известно разпределение на функциите (вж.
свързването на азот по-долу). Истинска многоклетъчност обаче при прокариотите не
се открива.
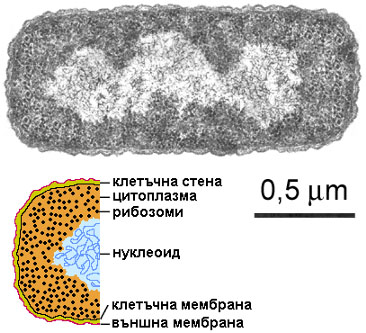
Накратко устройството на прокариотната клетка е следното: Тя
е оградена с липидна клетъчна мембрана. В средата на клетката се разполага
генетичният й апарат – една пръстенна молекула ДНК, често наричана бактериална
хромозома. Участъкът, зает от хромозомата, се нарича нуклеоид. Функционално той
е аналог на еукариотното ядро, но няма обвивка, оттам и името прокариоти.
Пространството между нуклеоида и клетъчната мембрана е изпълнено с цитоплазма,
съдържаща рибозоми. В нея може да има и извънхромозомна ДНК – малки пръстенни
молекули, наречени плазмиди.
Устройство на бактериална клетка –
електронно-микроскопска снимка на E. coli, любезно предоставена от Carl
Robinow, и обяснителна схема.
Някои прокариоти имат вътреклетъчни мембранни образувания, наречени ламели и тилакоиди. Те обикновено са вгъвания на клетъчната мембрана, по-рядко отделни мехурчета, и служат да увеличат повърхността й за нуждите на метаболизма. Срещат се предимно у автотрофните еубактерии и съдържат молекули-участници във фото- или хемосинтезата.
Външно от
клетъчната мембрана почти всички прокариоти имат клетъчна стена с опорна и
защитна функция. Еубактериалната клетъчна стена е изградена от дълги вериги
азотсъдържащ хетерополизахарид, омрежени с къси пептиди. Този сложен полизахарид
се нарича пептидогликан или муреин. Първият използван в медицината антибиотик,
пеницилинът, спира синтезата на муреин. Нашите телесни течности съдържат ензима
лизозим (мурамидаза), който разгражда муреина.
При еубактериите има два
основни типа строеж на клетъчната стена, които личат по цвета на клетката след
т. нар. оцветяване по Грам (Gram). Грам-положителните бактерии имат стена,
съдържаща до 40 слоя муреин и малко други съставки. При Грам-отрицателните
бактерии муреиновата мрежа е еднослойна, а над нея има липидна мембрана, подобна
на клетъчната, но много по-пропусклива. Нарича се външна мембрана. От нея навън
стърчат въглехидратните вериги на липополизахариди. Те, заедно със самата външна
мембрана, осигуряват на муреиновия слой известна защита срещу неблагоприятни
фактори (например лизозим).
3. Метаболизъм
Повечето
прокариоти са хетеротрофи – сапрофити и паразити. Част от тях имат обмяна на
веществата, подобна на нашата, т.е. освен че са хетеротрофи, те са и аероби.
Клетките им секретират навън смилателни ензими. Ако околната среда съдържа
биополимери, те се разграждат чрез хидролиза. Получените органични мономери
преминават през мембраната вътре в клетката. Някои от тях се включват в синтези.
Останалите, които ще се използват за енергия, се разграждат допълнително в
цитозола от ензимите на гликолизата и цикъла на Кребс. Отделените при
окислението електрони (НАД.Н) достигат до клетъчната мембрана. По вътрешната й
страна са разположени ензимите на дихателната верига, които пренасят електроните
до кислорода. Ензимът АТФ-синтетаза, който осъществява окислителното
фосфорилиране, е на същото място.
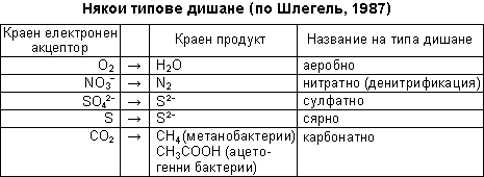
Освен описания тип обмяна на веществата
за прокариотите са известни много други. Например не всички дихателни вериги са
като нашите. Някои са по-къси и вместо с кислород завършват с друго вещество,
чиято редуцирана форма се отделя вместо вода. Процесът се нарича анаеробно
дишане. Например метанобактериите (гореспоменатите блатни архебактерии) дишат,
като редуцират СО2 до СН4. Те отговарят за отделянето на метан ("блатен газ") в
блатата. Сулфат-редуциращите бактерии дишат, превръщайки сулфатните йони и други
сравнително окислени форми на сярата в сероводород. Тяхна е "заслугата" за
отровените дълбочини на Черно море.
Редица прокариоти
изобщо нямат дихателни вериги. АТФ се синтезира само чрез прегрупиране на
атомите на съединенията, използвани като енергийни източници. Такива катаболитни
пътища се основават на гликолизата и общо се наричат ферментации. Според
основния си продукт ферментациите биват алкохолна, млечнокисела, пропионокисела,
мравченокисела, масленокисела и оцетнокисела.
Както ферментациите, така и
анаеробното дишане са свойствени на прокариоти, които могат да живеят и в
отсъствие на О2 (факултативни анаероби) или само в отсъствието му (строги или
облигатни анаероби)
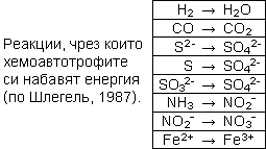
Макар че повечето прокариоти са хетеротрофи, има и
такива, които се хранят автотрофно. Цианобактериите фотосинтезират като зелените
растения. Други бактерии фотосинтезират, като вместо H2O използват H2S и вместо
O2 отделят S. И накрая, някои прокариоти са хемоавтотрофи – набавят си свободна
енергия чрез окисление на неорганични вещества, за което не им трябва светлина.
Например амоняк се окислява до нитрит, нитритът до нитрат, сероводород до
сулфат. Този автотрофен анаболизъм се нарича хемосинтеза.
Метаболитното
разнообразие на прокариотите е впечатляващо, особено ако го сравним с
еднообразието при еукариотите. Можем да заключим, че нашата обмяна на веществата
съвсем не е единственият съвместим с живота тип. Някои от прокариотните обменни
пътища, например ферментациите и фотосинтезата без отделяне на кислород,
изглеждат примитивни и вероятно са останки от ранната еволюция. Поне
хемосинтезата обаче трябва да е възникнала късно, защото изисква свободен
кислород. Следователно прокариотната клетъчна организация позволява радикални
промени в метаболизма и е добра основа за биохимична еволюция.
Важното за
цялата биосфера преобразуване на молекулния азот N2 до използваеми съединения
като NH3, наречено свързване или фиксиране на азота, се извършва изключително от
прокариоти. Някои от тях са сапрофити, обитаващи почвата или водите. Най-видният
им представител е почвената бактерия Azotobacter. Други живеят в симбиоза с
бобовите растения, като получават от растението органични вещества и в замяна му
осигуряват свързан азот. Наричат се грудкови бактерии, защото под тяхно влияние
коренът на бобовото растение образува грудки, в които те се заселват. Трета
важна група азот-свързващи прокариоти са нишковидните цианобактерии. Понеже
кислородът, отделен при фотосинтезата, пречи на свързването на азота, възниква
"разделение на труда". Повечето клетки от нишката са фотосинтезиращи, а тук-там
някоя се специализира за свързване на азот и се изхранва от съседите си.
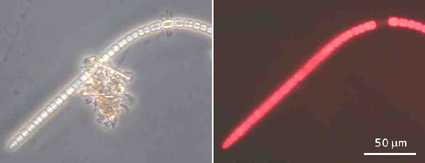
Неидентифицирана нишковидна цианобактерия, снимана при фазов
контраст (вляво) и автофлуоресценция (вдясно). Фотосинтетичните багрила при
осветяване със зелена светлина флуоресцират в червено. Една клетка остава тъмна,
защото е нагодена да свързва азот и не съдържа пигменти. До цианобактерията има
група бактерии, които не са фотосинтезиращи и затова не флуоресцират. От
Schauder (1997) с любезното разрешение на авторите Tom Lie и Rolf
Schauder.
4. Размножаване
При благоприятни условия прокариотната клетка непрекъснато расте и постоянно
синтезира ДНК. Периодично настъпва делене, което не смущава останалите жизнени
процеси. За целта получените при репликацията две бактериални хромозоми се
оттеглят в срещуположните краища на клетката, а в средата й клетъчната стена
нараства във вид на стесняваща се пръстенна бразда.
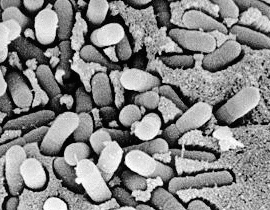
Електронно-микроскопска снимка на
деляща се сенна пръчица Bacillus subtilis в надлъжен срез. Оригинално увеличение
180 000 х. От Черепова (1989) с любезното й разрешение.

Прокариотите
могат да се размножават много бързо. При подходящи условия клетките им се делят
на всеки половин час.
Прокариотите не се размножават полово и нямат
истински полов процес. Най-близкото му при тях е т. нар. бактериална конюгация,
която ще бъде разгледана в друг раздел.
5. Осъществяване на наследствената информация и разпределяне на белтъците
Ако не се реплицират точно в момента, гените на прокариотите са
готови да служат като матрици за синтеза на РНК. Преди още молекулите мРНК да са
завършени и отделени от матрицата, върху предния им край се прикрепят рибозоми.
Доколкото за прокариотната клетка пространството се дели само на "вътре" и
"навън", съдбата на получените белтъци е проста. Те или остават във вътрешността
на клетката, или се включват в състава на мембраната, или се секретират. Когато
има външна мембрана, присъствието й не усложнява съществено разпределението на
клетъчните продукти, понеже тя е силно пропусклива.
6. Биологичен модел: Escherichia coli
Изясняването на
кой да е жизнен процес изисква огромна изследователска работа, която неизбежно
се върши от голям брой хора в продължение на много години. Ако при това всяка
лаборатория изследва различен обект, това би затруднило обобщаването на данните,
понеже резултати, получени за един вид, съвсем не е задължително да важат и за
друг. Ето защо при изследвания на основните жизнени процеси се предпочитат
няколко вида, признати от цялата биологична общност за основни обекти или
"модели". Те имат особености, които ги правят подходящи за лабораторна работа.
Наистина това важи и за много други видове и при избора е имало елемент на
случайност, но след като даден вид вече е станал основен обект, няма причини да
се заменя с друг. Някои от моделните организми са прокариоти. Най-важна измежду
тях е чревната бактерия (чревната пръчица) Escherichia coli.
Родът
Escherichia е наречен в чест на Теодор Ешерих (Escherich), който изолира
чревната бактерия за пръв път през 1885. Семейството й, Enterobacteriaceae,
обединява подвижни Грам-отрицателни пръчковидни бактерии, факултативни анаероби.
Те се отнасят към най-разпространената група Грам-отрицателни еубактерии,
наречена поради своето разнообразие Proteobacteria (от митичното същество
Протей, способно да мени образа си).
Ентеробактериите се движат чрез
около 10 камшичета, разположени на различни места по страничните повърхности на
клетката. В безкислородни условия получават АТФ чрез мравченокисела ферментация,
а някои, включително E. coli – и чрез анаеробно дишане. Както показва името на
семейството, то включва редица чревни обитатели. Освен Escherichia тук спада
например причинителят на чревни инфекции Salmonella.
Доколкото E. coli е
най-просто устроеният биологичен модел, можем да разглеждаме клетката й като
прототип на клетката изобщо. Доколкото животът на Земята през по-голямата част
от своето съществуване е бил изцяло прокариотен, това е най-малкото, което
дължим на прокариотите. По традиция обаче в учебната литература основното
описание на клетката се прави върху еукариотна клетка. Така ученикът или
студентът остава с мъгляво понятие за клетката като жизнена единица, защото
всяко украшение на еукариотите се възприема като абсолютно необходимо за живота.
Според мен трябва винаги да се започва с прокариотната клетка.
Електронно-микроскопска снимка и схема на чревна пръчица беше дадена по-горе.
Ето и подробна рисунка, показваща макромолекулните съставки.
Схема на
напречен срез от малка част от клетка на Escherichia coli. Клетъчната стена и
двете заграждащи я мембрани (външна и клетъчна) са зелени. Двойният липиден слой
на мембраните е натъпкан с мембранни белтъци, от външната стърчат и
липополизахариди. Показано е едно камшиче. В цитоплазмата се виждат виолетови
рибозоми, сини ензими и бели молекули мРНК. ДНК е жълта, на места свързана с
тъмножълти белтъци. Една оранжева ДНК-полимераза в момента извършва репликация.
От Goodsell (1999) с любезното му разрешение.
Наследственият материал (геномът) на
Escherichia coli е разчетен изцяло. Гените й са малко повече от 4000, т.е. почти
10 пъти по-малко, отколкото при бозайниците. Въпреки това метаболизмът на
чревната бактерия е много по-пълноценен от нашия. Ние сме лишени от редица
ензими, нужни за получаване на едни метаболити от други, и затова имаме твърде
сложни хранителни потребности. Храната трябва да ни осигури не само енергия и
необходимите химични елементи, а и голям брой конкретни съединения, които не
можем да синтезираме сами: незаменими аминокиселини, ненаситени мастни киселини,
витамини и др. Съвсем различно е при ентеробактериите. Добре развитите им
биохимични пътища ги правят невзискателни към храната. Достатъчно е тя да
съдържа глюкоза или друг въглехидрат за въглероден и енергиен източник, амоняк
за азотен източник и минерални соли. От тези съединения клетката синтезира
всички останали, които й трябват.
Обсъждайки обмяната на E. coli и изобщо
на ентеробактериите, трябва да споменем, че те могат да разграждат лактозата.
Този дизахарид е рядък в растителното царство и изобщо се среща в природата
главно в състава на млякото. Затова малко бактерии, най-вече обитатели на
храносмилателния път на бозайниците, могат да усвояват лактозата. Този белег се
оказва важен за молекулната биология; както ще видим по-късно, той преминава
през цялата й история като червена нишка.
Лесното отглеждане на E. coli е
една от причините да я изберат за основен прокариотен модел. Тя се размножава
бързо дори за прокариот: при благоприятни условия се дели на всеки 20 минути.
Макар нормално да живее в анаеробна среда при температура 37оС, тя понася и
използва кислорода и може да се развива и при 20-22оС.
При отглеждането
(култивирането) на бактерии хранителната среда или се налива в епруветки, както
е в течно състояние, или се разпределя в плитки и широки покрити съдове (т. нар.
Петриеви панички или петрита) и се желира. За целта в нея предварително се
добавя агар – извлек от определени водорасли от Индийския океан. Агарът съдържа
полизахарида агароза, който при стайна температура образува гел.
Когато
върху такъв гел се отглеждат бактерии, те не могат да се придвижват на дълги
разстояния. Потомците на всяка клетка остават близо един до друг и образуват
струпване, наречено колония (да не се бърка с колонията като група физически
свързани клетки). Ако в едно петри се посеят сравнително малък брой клетки,
колониите им ще бъдат отделни една от друга и след 24 часа ще се виждат с просто
око като изпъкнали кръгчета.
Петри с колонии E. coli. Диаметърът
на съда е 9-10 cm. Снимката е любезно предоставена от Стоян Чакъров от Софийския
университет.

Бактериите,
обитаващи червата ни, образуват значителна част (до 30%) от фекалната маса. След
като напуснат храносмилателния път, съдбата им е различна. Много от тях са
облигатни анаероби и загиват, ако попаднат в кислородна среда. Други се
размножават успешно в почва или вода и всъщност по-често се срещат там,
отколкото в храносмилателния път. Escherichia coli заема междинно място: оцелява
сравнително дълго във водните басейни, но не може да ги засели трайно. Затова
служи като показател за чистотата на питейната вода. Ако от даден водоизточник
може да се изолира E. coli, това значи, че е замърсен с фекалии.
По
правило E. coli е безвредна и дори полезна за човека. Заедно с другите обичайни
чревни бактерии тя снабдява своя гостоприемник с биотин и витамин К. Лечението с
антибиотици понякога води до хиповитаминоза по тези витамини. Освен това
ешерихията и другите нормални обитатели на храносмилателния път пречат на
заселването му с патогенни бактерии, понеже са техни успешни конкуренти. Има
обаче щамове Escherichia coli, които са ентеропатогенни или уропатогенни, т.е.
причиняват инфекции на храносмилателния или пикочния път. Ето снимка на такива
клетки, настанили се в чревния епител.
Ентеропатогенна E. coli
под сканиращ електронен микроскоп. Снимката е любезно предоставена от Stuart
Knutton от Бирмингамския университет.
7. Схващането за "минимален живот" и микоплазмите
Различните
организми и техните клетки силно се различават по сложност. Можем да се запитаме
как трябва да изглежда възможно най-простата жива система. Явно тя е малка
клетка, отделена от външната среда с липидна мембрана. За съхраняването на
наследствената информация е нужна молекула ДНК, а за разчитането й – рибозоми. В
цитоплазмата трябва да се остави място и за някои ензими, РНК и
метаболити.
Трудно е да се прецени умозрително колко гени са достатъчни
за живота. Известен е вирус само с 4 гена (бактериофагът QBeta), но тук
обсъждаме пълноценна жива система – клетка, която не разчита на чужди
биополимери. Явно са необходими рРНК и тРНК; рибозомни белтъци; белтъци,
осигуряващи деленето; ензими за репликацията, транскрипцията и транслацията,
както и за някакъв макар и най-прост метаболизъм; транспортни белтъци в
мембраната; вероятно и няколко регулаторни и структурни белтъка. По груби сметки
за самовъзпроизвеждащ се живот са нужни поне 250 гена. Молекула ДНК, достатъчно
дълга да ги кодира, заема обема на кълбо с диаметър 100 nm. Като се имат предвид
и рибозомите (диаметър около 20 nm), "минималната" жива система трябва да е
кълбо с диаметър около 200 nm. Метаболизмът на тази клетка ще бъде толкова
непълноценен, че трябва да си я представяме или като обитател на древната Земя,
където много реакции са протичали абиогенно, или като облигатен
паразит.
Най-малките и прости действителни организми, които са ни
известни, почти отговарят на горното описание. Това са група еубактерии,
наречени микоплазми. Те са облигатни паразити по различни животни и растения.
Някои от тях причиняват плевропневмонии и така групата е била открита
първоначално. Като патогени за цялостния организъм микоплазмите не са сред
най-опасните, но много успешно заразяват отглеждани в хранителна среда клетки от
бозайници и затова са голям проблем при клетъчното култивиране.
За
разлика от другите прокариоти микоплазмите нямат стена. Най-дребните измежду тях
са с естествената от физична гледна точка кълбовидна форма и диаметър 200 – 300
nm.
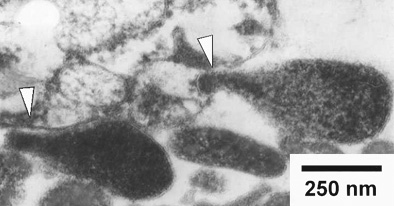
Електронно-микроскопска снимка на Mycoplasma genitalium,
любезно предоставена от Jens Blom и Jorgen Jensen от Статенс Серум Институт,
Копенхаген, Дания. Главичките на стрелки сочат места на прикрепване към
клетката-гостоприемник.
Геномът на
Mycoplasma genitalium, показана на горната снимка, е разчетен. Той съдържа 470
гена за белтъци, 3 за рРНК и 33 за тРНК, или общо почти 10 пъти по-малко,
отколкото при E. coli. Клетката си набавя АТФ чрез млечнокисела и оцетнокисела
ферментация. Редица важни биохимични пътища като синтези на аминокиселини,
нуклеотиди и мастни киселини практически липсват. Затова микоплазмите се
свързват плътно с повърхността на клетката-гостоприемник, както се вижда на
снимката, и извличат многобройните си необходими метаболити от нейната
цитоплазма. За разлика от вирусите микоплазмите все пак могат да се отглеждат и
без клетки-гостоприемници, но изискват среда със сложен състав поради твърде
бедния си метаболизъм.
8. Еубактерии и археи. Еволюционни връзки между прокариоти и еукариоти
Доколкото всички изучени форми на живот са принципно сходни, смята
се, че в далечното минало те са имали общ предшественик. Следователно всички
съвременни видове, както и измрелите, са в някакви родствени връзки помежду си.
При сравняване на организми винаги се интересуваме от еволюционното им родство.
Този въпрос е толкова по-сложен, колкото по-отдалечени са разглежданите групи.
Например трудно е да се установят точните родствени връзки между прокариотите и
еукариотите, понеже те са дивергирали (разделили са се в еволюцията си) много
отдавна. Това е логично предвид на големите разлики в устройството и
функционирането на техните клетки. По-неочаквано е, че между самите прокариоти
се очертават случаи на също толкова далечно родство. Например изглежда, че
еубактериите и архебактериите са се разделили още преди еукариотите да се
отцепят от прокариотите.
Архебактериите (археите) имат скромно
практическо значение, а за медицината – никакво, но поради необикновеното им
място в живия свят не бива да ги пренебрегваме. Клетъчната им стена е изградена
не от муреин, а от други полизахариди и от белтъци. Мембраната им е с необичаен
състав: липидите, които я изграждат, не съдържат мастни киселини. Камшичетата на
подвижните археи, макар да се въртят свободно като тези на бактериите и да им
приличат външно, имат различно устройство и състав. По метаболизма си археите
приличат на еубактериите, а по информационните си процеси (репликация,
транскрипция, транслация) – на еукариотите. Затова се предполага, че в
еволюцията първо са археите са се отделили от еубактериите, а след това от някоя
архея са произлезли еукариотите.
Тази представа е донякъде неочаквана,
понеже след като сме обединили всички прокариоти в една група, сме склонни
автоматично да я смятаме за монофилетична. Монофилетична се нарича група, която
включва всички потомци на един вид-предшественик. Група, която включва някои, но
не всички потомци на даден предшественик, се нарича парафилетична, а група,
обединяваща два или повече независими монофилетични таксона –
полифилетична.
Още от времето на Дарвин, според когото класификацията на
живия свят трябва да е негово родословно дърво, има стремеж видовете да се
подреждат в монофилетични групи. Това обаче, освен че често е трудно, понякога
не е и уместно. Прокариотите си приличат в много отношения и е удобно да се
разглеждат заедно. Няма обаче основания да ги смятаме за монофилетична група,
понеже всичките им общи белези изглеждат примитивни. Общите белези са надежден
знак за близко родство само ако са апоморфни. Примитивните не са показателни,
защото могат да се запазят у две групи неограничено дълго след дивергенцията им
от общия предшественик. Най-вероятно точно такива са приликите между еубактерии
и архебактерии. Следователно групата Прокариоти е парафилетична, понеже има общ
предшественик (предполагаемия прародител на всички организми на Земята), но
изключва някои негови потомци, а именно еукариотите.
В края на главата за
прокариотите е уместно да оборим някои предразсъдъци спрямо тях. Сравнени с
неживите тела, дори и микоплазмите са крайно високо организирани. Ако обаче
човек е свикнал да мисли за еукариотната клетка, простотата на прокариотите може
да не му хареса. Доколкото първите организми не може да са били други освен
прости, а еволюцията често е свързана с усложняване, ние сме склонни да
подценяваме по-простите форми на живот. Случва се без други основания да ги
приемаме не само за примитивни, а и за зле адаптирани и биологично непълноценни.
Често се използва понятието "низши", което няма място в съвременната биология.
Всъщност еволюцията често протича със запазване на простата организация, а
понякога дори с вторично опростяване. Това допуска отлична адаптация. Само
естественият отбор позволява, и то с уговорки, да степенуваме организмите по
биологични качества. От тази гледна точка всички видове, които сега или в
произволен избран момент още не са измрели, са еднакво
"висши".
Примитивните структури са генерализирани, т.е. способни са да
изпълняват доста функции, без да са добре нагодени за никоя от тях. Понякога в
еволюцията структурите се видоизменят така, че да изпълняват една функция
възможно най-добре. Тогава те се наричат специализирани. Генерализираните форми
могат да еволюират в много посоки, а специализацията е свързана с избор на
определена посока.
Примитивната клетка е била малка, просто устроена и
генерализирана. Съвременните прокариоти също са малки и просто устроени, но
простотата им в някои отношения е толкова рационална, че може да се обясни само
със специализация. И в наследствения материал на бактериите, и във фенотипа им е
трудно да се открие нещо излишно. Прокариотната клетъчна организация, при която
всички жизнени процеси протичат едновременно и на едно място, осигурява тяхната
бързина и съгласуваност. В резултат прокариотите се приспособяват към промените
в средата с немислима за еукариотите ефективност. Достатъчно е да се посочи, че
при идеални условия типичната прокариотна клетка се удвоява за 30 минути, а
типичната еукариотна – за 24 часа. Следователно прокариотите не са по-низши от
еукариотите, а са възприели друга стратегия на адаптация. Заради ефективността
те са пожертвали заложбите си за по-висока организация.


Коментари